|


 Curently online :
8 Curently online :
8 |
|
 Total visitors :
7515928 Total visitors :
7515928 |
|
|
Mass spider silk production through targeted gene replacement in Bombyx mori
Monday, 2018/08/27 | 09:09:47
|
|
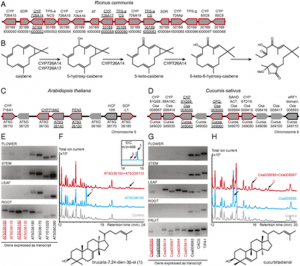
Jun Xu, Qinglin Dong, Ye Yu, Baolong Niu, Dongfeng Ji, Muwang Li, Yongping Huang, Xin Chen, and Anjiang Tan PNAS August 6, 2018. SignificanceThe use of heterologous systems to express spider silk has become an attractive method. However, achieving cost-effective production and high yields is still challenging. Here, we describe the establishment of a targeted gene replacement system in Bombyx mori to express the major ampullate spidroin-1 gene (MaSp1) from the spider Nephila clavipes. With the aid of transcription activator-like effector nuclease-mediated homology-directed repair, we genetically replaced the silkworm fibroin heavy chain gene with MaSp1 with considerable transformation efficiency, and the chimeric MaSp1 yields reached up to 35.2% wt/wt of cocoon shells in transformed silkworms. The genetically modified silk fiber had significant changes in mechanical properties, with improved extensibility. This system will shed light on the future mass production of new biomaterials, including spider silk. AbstractSpider silk is one of the best natural fibers and has superior mechanical properties. However, the large-scale harvesting of spider silk by rearing spiders is not feasible, due to their territorial and cannibalistic behaviors. The silkworm, Bombyx mori, has been the most well known silk producer for thousands of years and has been considered an ideal bioreactor for producing exogenous proteins, including spider silk. Previous attempts using transposon-mediated transgenic silkworms to produce spider silk could not achieve efficient yields, due to variable promoter activities and endogenous silk fibroin protein expression. Here, we report a massive spider silk production system in B. mori by using transcription activator-like effector nuclease-mediated homology-directed repair to replace the silkworm fibroin heavy chain gene (FibH) with the major ampullate spidroin-1 gene (MaSp1) in the spider Nephila clavipes. We successfully replaced the ∼16-kb endogenous FibH gene with a 1.6-kb MaSp1 gene fused with a 1.1-kb partial FibH sequence and achieved up to 35.2% chimeric MaSp1 protein amounts in transformed cocoon shells. The presence of the MaSp1 peptide significantly changed the mechanical characteristics of the silk fiber, especially the extensibility. Our study provides a native promoter-driven, highly efficient system for expressing the heterologous spider silk gene instead of the transposon-based, random insertion of the spider gene into the silkworm genome. Targeted MaSp1 integration into silkworm silk glands provides a paradigm for the large-scale production of spider silk protein with genetically modified silkworms, and this approach will shed light on developing new biomaterials.
See: http://www.pnas.org/content/early/2018/07/31/1806805115
Figure 1: Schematic representation of the TALEN-mediated gene replacement system and the targeted integration of transgene constructs. (A) Schematic representation of the FibH gene and the TALEN targeting sites. The thin yellow line represents the FibH genomic locus, with the open boxes signifying the promoter, exons, and poly(A) signal. A 1,166-bp fragment (green box) located at the 5′ end represents the promoter region. The two 42-bp and 15,750-bp fragments (pink boxes) are exon 1 and exon 2, respectively. A 300-bp fragment (gray box) located at the 3′ end is the poly(A) signal. The red and green lightning icons indicate the two TALEN target sites. In the PJET-Red and PJET-MaSp1 donor constructs, a DsRed2 marker expression cassette driven by a baculovirus IE1 promoter was cloned into the PJET-1.2 vector. DNA fragments of 1,356 bp and 1,331 bp at the 5′ and 3′ ends flanking the TALEN sites were PCR-amplified, subcloned into vectors, and used as homologous arms (L-homo and R-homo, respectively). The 1,593-bp MaSp1 partial sequence is shown by the green box. Primer positions for amplification analyses of the integrated insertions in transformed silkworms are shown by arrows. Primer pairs of F1/R1 (or R3) and F2/R2 were used to amplify the 5′- and 3′-end insertion junctions, respectively. (B) Sequencing results of the integrated diagnostic DNA fragments to show 5′ and 3′ junction genome–donor integration. |
|
|
|
[ Other News ]___________________________________________________
|
(38).png "Mass spider silk production through targeted gene replacement in Bombyx mori")