|


 Curently online :
19 Curently online :
19 |
|
 Total visitors :
7660309 Total visitors :
7660309 |
|
|
The evolving battle between yellow rust and wheat: implications for global food security
Saturday, 2022/04/02 | 08:28:11
|
|
Laura Bouvet, Sarah Holdgate, Lucy James, Jane Thomas, Ian J. Mackay & James Cockram Theoretical and Applied Genetics; March 2022; vol. 135: 741–753 AbstractWheat (Triticum aestivum L.) is a global commodity, and its production is a key component underpinning worldwide food security. Yellow rust, also known as stripe rust, is a wheat disease caused by the fungus Puccinia striiformis Westend f. sp. tritici (Pst), and results in yield losses in most wheat growing areas. Recently, the rapid global spread of genetically diverse sexually derived Pst races, which have now largely replaced the previous clonally propagated slowly evolving endemic populations, has resulted in further challenges for the protection of global wheat yields. However, advances in the application of genomics approaches, in both the host and pathogen, combined with classical genetic approaches, pathogen and disease monitoring, provide resources to help increase the rate of genetic gain for yellow rust resistance via wheat breeding while reducing the carbon footprint of the crop. Here we review key elements in the evolving battle between the pathogen and host, with a focus on solutions to help protect future wheat production from this globally important disease.
See: https://link.springer.com/article/10.1007/s00122-021-03983-z
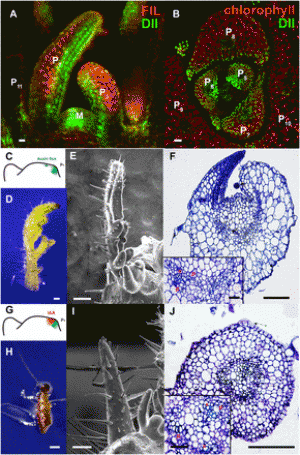
Figure 1: The Puccinia striiformis Westend f. sp. tritici (Pst) lifecycle. Wheat Pst infection at the adult plant stage in wheat, showing a yellow, and b orange coloured pustules that shed urediniospores. c Symptoms of Pst infection of the wheat ear. False colour scanning electron microscopy images of d urediniospores, e pustules on the wheat leaf surface, and f pustule in cross section. g Diagram illustrating the main features of the Pst lifecycle. Top left: Wheat plants can be infected by, (i) wind-blown single-cell dihaploid dikaryotic urediniospores (containing one haploid genome copy in each of the two nuclei within the cell: n + n′) produced on wheat, or (ii) by aeciospores (n + n′) produced on the alternative host (Berberis spp.). Yellow rust infection is typically observable on the heat upper leaf surface as parallel rows of yellow to orange pustules which release urediniospores, resulting in cycles of re-infection and cross-infection in wheat. Top middle: at ear emergence, yellow rust infection can occur on the florets of the wheat ear. Top right: towards the end of the wheat season, telia may form on the underside of the epidermis, from which diploid doubled haploid (2*n) two-celled teliospores are produced by karyogamy. Teliospores readily germinate to produce haploid basidiospores. Bottom right: Basidiospores germinate and infect leaves of the alternate host. Bottom middle: Basidiospore infection leads to the production of pycnia, typically on the upper side of the leaves, which release haploid pycniospores. Fusion of pycniospores with the receptive hypha of a mating-type compatible pycnia leads to dikaryotization and the development of aecia on the leaf underside. Bottom left: Aecia release vegetative aeciospores (n + n′) which are only able to infect the primary host species (predominantly wheat). For more information, see Chen et al. (2014), Schwessinger (2016) or Mehmood et al. (2020) (colour figure online). |
|
|
|
[ Other News ]___________________________________________________
|
(66).png "The evolving battle between yellow rust and wheat: implications for global food security")