|


 Đang trực tuyến :
36 Đang trực tuyến :
36 |
|
 Số lượt truy cập :
34063872 Số lượt truy cập :
34063872 |
|

|

|

|
|
Đa dạng di truyền xét trên trình tự gen của loài sắn hoang dại và giống sắn trồng.
Thứ hai, 30-05-2016 | 09:11:53
|
|
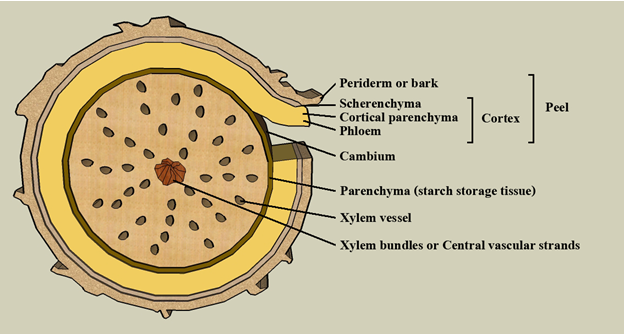
SẮN (Manihot esculenta) cung cấp calories và dinh dưỡng cho hơn nửa tỷ người trên thế giới. Sắn đã được con người ở vùng Amazone thuần hóa từ loài hoang dại sang loài sắn trồng thông qua việc trồng trọt loài tổ tiên M. esculenta ssp. Flabellifolia. Hiện nay, giống sắn trồng được canh tác trên vùng nhiệt đới và trên toàn thế giới. Bredeson và ctv. (2016) tạo nên một bộ genome tổng hợp chất lượng cao của cây sắn với sự tiếp giáp lẫn nhau được cải thiện rất tốt (contiguity), sự liên kết được cải thiện, cũng như tính chất hoàn chỉnh của bộ genome; có khoảng 97% gen được neo (anchored) thành công trên từng nhiễm sắc thể. Bredeson và ctv. (2016) thấy rằng độ tứ bội thể có tính chất “paleotetraploidy” của cây sắn là kết quả có thể chia sẻ thông tin với genome của cây cao su (Hevea), đó là nguồn cơ sở dữ liệu mang tính chất so sánh rất tốt. Họ tiến hành giải trình tự của tập đoàn giống sắn được thu thập trên toàn thế giới với 58 mẫu giống sắn, bao gồm giống trồng trọt và mẫu hoang dại có l;iên quan gần với giống trồng ví dụ nhự mẫu Ceará hoặc India rubber (M. glaziovii), và 268 mẫu giống sắn trồng của Châu Phi. Có một sự dàn trải khá rộng mang tính chất “admixture” giữa các loài. Họ phát hiện dấu vết di truyền (genetic signature) của những chương trình lai tạo giống sắn của quá khứ. Sắn là loài cây trồng nhân giống vô tính bằng hom thân, sắn cũng là loài có thể bị tổn thương do nguồn gây bệnh (pathogens) cũng như stress phi sinh học. Nguồn cơ sở dữ liệu có tính chất genomic như vậy có thể sẽ là thông tin hữu ích cho tương lai giúp việc cải tiến gióng sắn thành công. CHỈ THỊ PHÂN TỬ IRAP & REMAPRetrotransposons vô cùng phong phú trong nhiều genome cây trồng trong đó có bộ genome của cây sắn. Kuhn và ctv. (2016) đã phát triển bộ chỉ thị IRAP (inter-retrotransposon amplified polymorphism) và chỉ thị REMAP (retrotransposon-microsatellite amplified polymorphism) để phục vụ nghiên cứu bộ genome cây sắn (Manihot esculenta Crantz). Bốn giống sắn được xem xét là Fécula Branca, IPR-União, Olho Junto, và Tamboara, mỗi giống được thu hai mẫu phục vụ khảo nghiệm đánh dấu vân tay bằng marker (IRAP and REMAP fingerprints). Mười hai cặp mồi đã được thiết kế và cho khuếch đại mẫu theo hai nghiệm thức: (1) riêng biệt nhau; và (2) phối trộn nhau. Có 42 tổ hợp IRAP/REMAP primer đã khuếch đại được 431 đoạn phân tử DNA nghiên cứu (số băng; số marker) trong đó người ta thấy 36 phân tử (8.36%) biểu hiện đa hình. Số chỉ thị có thông tin lớn nhất là 16 markers được tìm thấy với các primers AYF2 và AYF2 x AYF4. Số băng của mỗi primer biến thiên từ 3 đến 6 băng, với giá trị trung bình là 10,26 đoạn phân tử được khuếch đại tương ứng với mỗi primer. Kích thức của sản phẩm PCR biến thiên 100 - 7000 bp. Mồi AYF2 phát sinh số lượng lớn nhất đoạn phân tử được khuếch đại và có kết quả số băng đa hình cao nhất (68.75%). Hai mẫu của mỗi giống sắn được người ta sử dụng để minh chứng tính chất hữu dụng và tính chất đa hình chủa chỉ thị phân tử IRAP/REMAP. IRAP và REMAP markers có số băng mang tính chất “reproducible” rất tốt, có thông tin di truyền và đáng tin cậy trong nghiên cứu đa dạng di truyền cũng như mối quan hệ huyết thống giữa các giống sắn (Kuhn và ctv. 2016). HỌ GEN WRKY CỦA GENOME SẮN LIÊN QUAN ĐẾN CHỐNG CHỊU HẠNHọ gen WRKY là một họ rất lớn mã hóa protein là những yếu tố phiên mã (TFs) có trong thực vật bậc cao, đóng vai trò trung tâm của những tiến trình sinh lý và thích ứng với môi trường. Tuy nhiên, hiện có rất ít thông tin về họ gen WRKY trong bộ genome cây sắn (Manihot esculenta). Wei và ctv. (2016) đã phân lập được 85 gen của WRKY trong genome cây sắn. Họ chia chúng ra thành 3 nhóm di truyền khác nhau tùy theo những domain WRKY có tính chất bảo thủ và tùy theio kiến trúc “zinc-finger”. Phân tích motif có tính chất bảo thủ ấy cho thấy rằng: tất cả những phân tử MeWRKYs đã xác định đều có chứa vùng domain WRKY rất bảo thủ. Phân tích kiến trúc gen cho thấy số introns của những gen MeWRKY biến thiên từ 1 đến 5, với bản chất các gen MeWRKY có 3 exons. Phổ thể hiện gen MeWRKY trong các mô khác nhau như vậy và trong phản ứng với stress khô hạn được các tác giả phân tích bằng kỹ thuật RNA-seq. Wei và ctv. (2016) thấy rằng có72 gen MeWRKY biểu hiện hết sức khác nhau trong những phân tử transcript vô cùng đa dạng ấy. Có 78 gen MeWRKY thể hiện khác biệt nhau khi đáp ứng với stress khô hạn trong những mẫu giống sắn trồng đã thu thập. Điều này chứng minh rằng chúng thật sự tham gia vào những tiến trình phát triển cây trồng, đặc biệt tính chống chịu hạn của cây sắn. Sau cùng, sự thể hiện của 9 gen WRKY đã được nhóm tác giả phân tích bằng qRT-PCR trong nghiệm thức quan sát áp suất thẩm thấu, nghiệm thức mặn, nghiệm thức ABA, H2O2, và nghiệm thức lạnh. Kết quả cho thấy những gen MeWRKYs có thể hiện diện trong các chu trình truyền tín hiệu khác nhau (Wei và ctv. 2016).
Hình: Phân tích di truyền huyết thống các protein WRKY của sắn và Arabidopsis. Cây gia hệ NJ được hình thành bởi những trình tự vùng domain của WRKY trong cây sắn và Arabidopsis thông qua phần mềm ClustalX 2.0 và MEGA5 với 1.000 bootstrap. Những nhành có ít hơn 50% bootstrap đều bị gãy. Những protein WRKY được xếp thành 3 nhóm chính (1–3) và 5 nhóm phụ (2a–2e). Nhóm protein 1có suffix “N” hoặc “C” chỉ ra gốc N trong những domain của WRKY hoặc gốc C của WRKY domains. “AtWRKYs” là những protein của Arabidopsis. “MeWRYKs” là những protein của sắn. GEN KHÁNG BỆNH THÁN THƯ
Tài liệu tham khảo
Utsumi Y, Tanaka M, Kurotani A, Yoshida T, Mochida K, Matsui A, Ishitani M, Sraphet S, Whankaew S, Asvarak T, Narangajavana J, Triwitayakorn K, Sakurai T, Seki M. 2016. Cassava (Manihot esculenta) transcriptome analysis in response to infection by the fungus Colletotrichum gloeosporioides using an oligonucleotide-DNA microarray. J Plant Res. 2016 May 2. http://link.springer.com/article/10.1007%2Fs10265-016-0828-x .
Kuhn BC, Mangolin CA, Souto ER, Vicient CM, Machado MF. 2016. Development of retrotransposon-based markers IRAP and REMAP for cassava (Manihot esculenta). Genet Mol Res. 2016 Apr 7;15(2). doi: 10.4238/gmr.15027149.
Bredeson JV, Lyons JB, Prochnik SE, Wu GA, Ha CM, Edsinger-Gonzales E, Grimwood J, Schmutz J, Rabbi IY, Egesi C, Nauluvula P, Lebot V, Ndunguru J, Mkamilo G, Bart RS, Setter TL, Gleadow RM, Kulakow P, Ferguson ME, Rounsley S, Rokhsar DS. 2016. Sequencing wild and cultivated cassava and related species reveals extensive interspecific hybridization and genetic diversity. Nat Biotechnol. 2016 May;34(5):562-70..
Wei Y, Shi H, Xia Z, Tie W, Ding Z, Yan Y, Wang W, Hu W, Li K. 2016. Genome-Wide Identification and Expression Analysis of the WRKY Gene Family in Cassava. Front Plant Sci. 2016 Feb 5;7:25. |
|
|
|
[ Tin tức liên quan ]___________________________________________________
|
(31).png "Đa dạng di truyền xét trên trình tự gen của loài sắn hoang dại và giống sắn trồng.")
(32).png "Đa dạng di truyền xét trên trình tự gen của loài sắn hoang dại và giống sắn trồng.") Bệnh thán thư trên khoai mì (sắn) được biết với thuật ngữ CAD (cassava anthracnose disease), do nấm Colletotrichum gloeosporioides f. sp. manihotis gây ra. Đây là bệnh khá nghiêm trọng cho cây khoai mì (Manihot esculenta) trên toàn thế giới. Utsumi và ctv. (2016) đã xây dựng quy trình chạy microarray với bộ “oligonucleotide-DNA” đại diện cho 59.079 probes tương ứng với khoảng 30.000 gen trên cơ sở dữ liệu của các phân tử đánh dấu trình tự gốc (original expressed sequence tags) và cơ sở dữ liệu chuỗi trình tự RNA của khoai mì. Người ta áp dụng thông tin này để nghiên cứu cơ chế tính kháng bệnh ở mức độ phân tử đối với sự xâm nhiễm của vi nấm trên hai giống khoai mì: Huay Bong 60 (HB60, kháng CAD) và giống Hanatee (HN, nhiễm CAD). Kết quả phân tích “quantitative real-time reverse transcription PCR” và kết quả phân tích phổ thể hiện bằng kỹ thuật microarray cho thấy rằng: sự thể hiện của những gen liên quan đến chức năng tự vệ của cây, ví dụ như họ gen PR (pathogenesis-related genes), họ gen liên quan đến thành tế bào, gen mã hóa enzyme có chức năng giải độc (detoxification), các gen có liên quan đến đáp ứng của cây đối với vi khuẩn, gen mã hóa MAPK (mitogen-activated protein kinase), các gen liên quan đến chu trình salicylic acid, jasmonic acid và ethylene với số lượng cao hơn rất nhiều trong giống khoai mì HB60 so với giống HN. Kết quả còn cho thấy sự kích hoạt họ gen PR trong giống HB60 khi có vi nấm xâm nhiễm vào cây, và sự thể hiện cao hơn của những gen liên quan đến chức năng tự vệ trong giống khoai mì HB60 so với giống nhiễm HN chứng tỏ giống HB60 có nguồn gen kháng bệnh thật sự. Kết quả cho thấy việc sử dụng bộ chip sinh học “cassava oligo microarray” có thể cải tiến được những hiểu biết của chúng ta về cơ chế sinh học phân tử có liên quan đến phản ứng của cấy đối với môi trường và sự phát triển. Lợi ích của chọn giống bằng kỹ thuật phân tử tỏ ra hữu dụng trong trường hợp cây khoai mì (sắn).
Bệnh thán thư trên khoai mì (sắn) được biết với thuật ngữ CAD (cassava anthracnose disease), do nấm Colletotrichum gloeosporioides f. sp. manihotis gây ra. Đây là bệnh khá nghiêm trọng cho cây khoai mì (Manihot esculenta) trên toàn thế giới. Utsumi và ctv. (2016) đã xây dựng quy trình chạy microarray với bộ “oligonucleotide-DNA” đại diện cho 59.079 probes tương ứng với khoảng 30.000 gen trên cơ sở dữ liệu của các phân tử đánh dấu trình tự gốc (original expressed sequence tags) và cơ sở dữ liệu chuỗi trình tự RNA của khoai mì. Người ta áp dụng thông tin này để nghiên cứu cơ chế tính kháng bệnh ở mức độ phân tử đối với sự xâm nhiễm của vi nấm trên hai giống khoai mì: Huay Bong 60 (HB60, kháng CAD) và giống Hanatee (HN, nhiễm CAD). Kết quả phân tích “quantitative real-time reverse transcription PCR” và kết quả phân tích phổ thể hiện bằng kỹ thuật microarray cho thấy rằng: sự thể hiện của những gen liên quan đến chức năng tự vệ của cây, ví dụ như họ gen PR (pathogenesis-related genes), họ gen liên quan đến thành tế bào, gen mã hóa enzyme có chức năng giải độc (detoxification), các gen có liên quan đến đáp ứng của cây đối với vi khuẩn, gen mã hóa MAPK (mitogen-activated protein kinase), các gen liên quan đến chu trình salicylic acid, jasmonic acid và ethylene với số lượng cao hơn rất nhiều trong giống khoai mì HB60 so với giống HN. Kết quả còn cho thấy sự kích hoạt họ gen PR trong giống HB60 khi có vi nấm xâm nhiễm vào cây, và sự thể hiện cao hơn của những gen liên quan đến chức năng tự vệ trong giống khoai mì HB60 so với giống nhiễm HN chứng tỏ giống HB60 có nguồn gen kháng bệnh thật sự. Kết quả cho thấy việc sử dụng bộ chip sinh học “cassava oligo microarray” có thể cải tiến được những hiểu biết của chúng ta về cơ chế sinh học phân tử có liên quan đến phản ứng của cấy đối với môi trường và sự phát triển. Lợi ích của chọn giống bằng kỹ thuật phân tử tỏ ra hữu dụng trong trường hợp cây khoai mì (sắn).